.svg)





What fisetin actually does at the cellular level
Fisetin is a flavonoid, a class of polyphenolic compounds found in plants. What distinguishes fisetin from other flavonoids is its classification as a senolytic, a compound that selectively induces death in senescent cells. Senescent cells are cells that have stopped dividing but resist the normal process of programmed cell death (apoptosis). They accumulate with age and secrete inflammatory molecules, growth factors, and proteases collectively known as the senescence-associated secretory phenotype (SASP). This secretory profile drives chronic low-grade inflammation, tissue dysfunction, and accelerates multiple hallmarks of aging.
Fisetin targets senescent cells by interfering with the BCL-2 family of proteins, which regulate the intrinsic mitochondrial apoptosis pathway. Senescent cells upregulate anti-apoptotic proteins like BCL-2, BCL-XL, and BCL-W to resist clearance. These proteins block the release of cytochrome c from mitochondria, preventing the activation of caspases that execute cell death. Fisetin disrupts this protective mechanism, allowing senescent cells to undergo apoptosis while leaving healthy proliferating cells largely unaffected. The selectivity is not absolute, but the threshold for triggering apoptosis is lower in senescent cells due to their reliance on these survival pathways.
The compound also modulates other pathways relevant to cellular aging:
- It activates sirtuins, particularly SIRT1, which influence DNA repair, mitochondrial function, and metabolic regulation.
- Fisetin reduces oxidative stress by upregulating antioxidant enzymes through the NRF2 pathway.
- In animal models, it improves markers of mitochondrial health and reduces markers of systemic inflammation.
These effects are mechanistically distinct from its senolytic action but may contribute to its overall impact on healthspan.
How fisetin connects to the hallmarks of aging
Fisetin's primary relevance to longevity lies in its ability to reduce the burden of cellular senescence, one of the established hallmarks of aging. Senescent cell accumulation is not just a marker of aging but a driver of it. These cells contribute to tissue dysfunction by secreting inflammatory cytokines like IL-6 and IL-8, matrix metalloproteinases that degrade extracellular structures, and growth factors that promote fibrosis. By clearing senescent cells, fisetin attenuates this cascade.
The compound also intersects with chronic inflammation (inflammaging). The SASP secreted by senescent cells is a major contributor to the systemic inflammatory tone that rises with age. Reducing senescent cell numbers through senolytic intervention lowers circulating inflammatory markers in animal models. This has downstream effects on multiple age-related pathologies, including cardiovascular disease, neurodegenerative conditions, and metabolic dysfunction.
Fisetin's effects on mitochondrial function connect it to another hallmark: mitochondrial dysfunction. Senescent cells often exhibit impaired mitochondrial respiration and increased production of reactive oxygen species. By clearing these cells and supporting mitochondrial biogenesis through SIRT1 activation, fisetin may help preserve cellular energy production. The compound also influences nutrient sensing pathways, particularly mTOR and AMPK, which regulate cellular growth, autophagy, and metabolic adaptation.
What drives senescent cell accumulation and how fisetin intervenes
Senescent cells accumulate for several reasons. DNA damage from oxidative stress, telomere attrition, oncogene activation, and mitochondrial dysfunction can all trigger the senescence program. The body has mechanisms to clear these cells through immune surveillance, primarily via natural killer cells and macrophages. But immune function declines with age, and the clearance rate falls behind the rate of senescent cell formation. The result is a net accumulation that accelerates tissue aging.
Fisetin intervenes by lowering the threshold for apoptosis in cells that have already entered senescence. It does not prevent cells from becoming senescent in the first place, nor does it reverse the senescent state. Instead, it selectively eliminates cells that are already contributing to tissue dysfunction. This is mechanistically different from senostatic agents (which suppress the SASP without killing the cells) or from interventions that prevent senescence induction altogether.
The effectiveness of fisetin as a senolytic varies by cell type and the trigger that induced senescence:
- In vitro studies show that fisetin is more effective at clearing senescent human umbilical vein endothelial cells than senescent fibroblasts.
- This variability reflects differences in the expression of BCL-2 family proteins and other survival pathways across cell types.
- In animal models, fisetin reduces senescent cell markers in adipose tissue, kidney, and heart, but the magnitude of the effect differs by tissue.
- This suggests that a single senolytic agent may not uniformly clear senescent cells across all tissues, and combination approaches may be necessary.
Dietary and environmental factors that influence senescent cell burden
Chronic caloric excess, particularly diets high in refined carbohydrates and saturated fats, accelerates senescent cell accumulation through metabolic stress and insulin resistance. Conversely, caloric restriction and time-restricted feeding reduce senescent cell burden in animal models, likely through enhanced autophagy and improved mitochondrial function. Physical inactivity also contributes; exercise activates pathways that promote senescent cell clearance and reduces systemic inflammation. Exposure to UV radiation, air pollution, and heavy metals increases DNA damage and oxidative stress, both of which trigger cellular senescence. Smoking is a particularly potent driver of senescent cell accumulation in lung tissue. Chronic psychological stress elevates cortisol, which has been linked to accelerated epigenetic aging and increased senescent cell markers in some tissues.
Why the same senolytic produces different outcomes in different people
Individual responses to fisetin are shaped by genetic variation, baseline senescent cell burden, and metabolic phenotype. Polymorphisms in genes encoding BCL-2 family proteins influence how readily senescent cells undergo apoptosis in response to senolytic agents. Individuals with higher baseline expression of anti-apoptotic proteins may require higher doses or combination therapies to achieve meaningful senescent cell clearance.
Baseline senescent cell burden varies widely even among individuals of the same chronological age. This reflects differences in cumulative exposure to stressors, immune function, and the efficiency of endogenous clearance mechanisms. Someone with a high senescent cell burden may experience more pronounced benefits from senolytic intervention, while someone with a low burden may see minimal change. Epigenetic clocks and senescence-associated biomarkers like p16INK4a can provide some indication of baseline burden, though these are not yet standard clinical tools.
Metabolic health also matters:
- Insulin resistance and chronic inflammation create a tissue environment that favors senescent cell survival and amplifies the SASP.
- Individuals with metabolic syndrome may have a higher proportion of senescent cells in adipose tissue and liver, and these cells may be more resistant to clearance.
- Conversely, individuals with better metabolic health may have lower baseline senescent cell numbers and more efficient immune-mediated clearance, potentially reducing the incremental benefit of exogenous senolytics.
Gut microbiome composition and senolytic metabolism
The gut microbiome influences the metabolism and bioavailability of flavonoids, including fisetin. Certain bacterial species can cleave glycosidic bonds and modify flavonoid structures, affecting absorption and systemic exposure. Individuals with dysbiotic microbiomes may metabolize fisetin differently, leading to variable plasma concentrations and tissue distribution. This is an emerging area of research, but it suggests that microbiome composition could be a source of inter-individual variability in senolytic efficacy.
What the research actually supports and where the evidence gets thinner
Animal studies provide the strongest evidence for fisetin's senolytic effects. In aged mice, intermittent fisetin treatment extends median and maximum lifespan, reduces senescent cell markers in multiple tissues, and improves physical function. The doses used in these studies (typically 100 mg/kg body weight administered intermittently) are far higher than what could be achieved through diet alone. For a 70 kg human, this would translate to roughly 7 grams of fisetin per dose, whereas a cup of strawberries contains approximately 0.16 milligrams.
Human clinical trials are limited in number and scope. A 2019 pilot study in women with obesity tested fisetin supplementation at doses of 100 mg twice daily for two days. The study found the intervention to be safe and feasible, with some reduction in inflammatory markers, but it was not designed to assess longevity outcomes. Other ongoing trials are evaluating fisetin in the context of osteoarthritis, chronic kidney disease, and frailty, but results are preliminary. The challenge is that senescent cell burden is difficult to measure non-invasively in humans, and surrogate markers like circulating inflammatory cytokines are influenced by many factors beyond senescence.
Bioavailability remains a significant limitation. Oral fisetin has poor absorption and rapid metabolism, resulting in low plasma concentrations. Novel formulations using liposomal encapsulation or nanoparticle delivery are being developed to address this, but these are not yet widely available. The distinction between senolytic and senostatic effects also matters. Some of fisetin's benefits in animal models may come from its antioxidant and anti-inflammatory properties rather than from direct senescent cell clearance. Disentangling these mechanisms in humans will require studies that measure senescent cell markers directly in tissue biopsies before and after treatment, which is invasive and not commonly done outside of research settings.
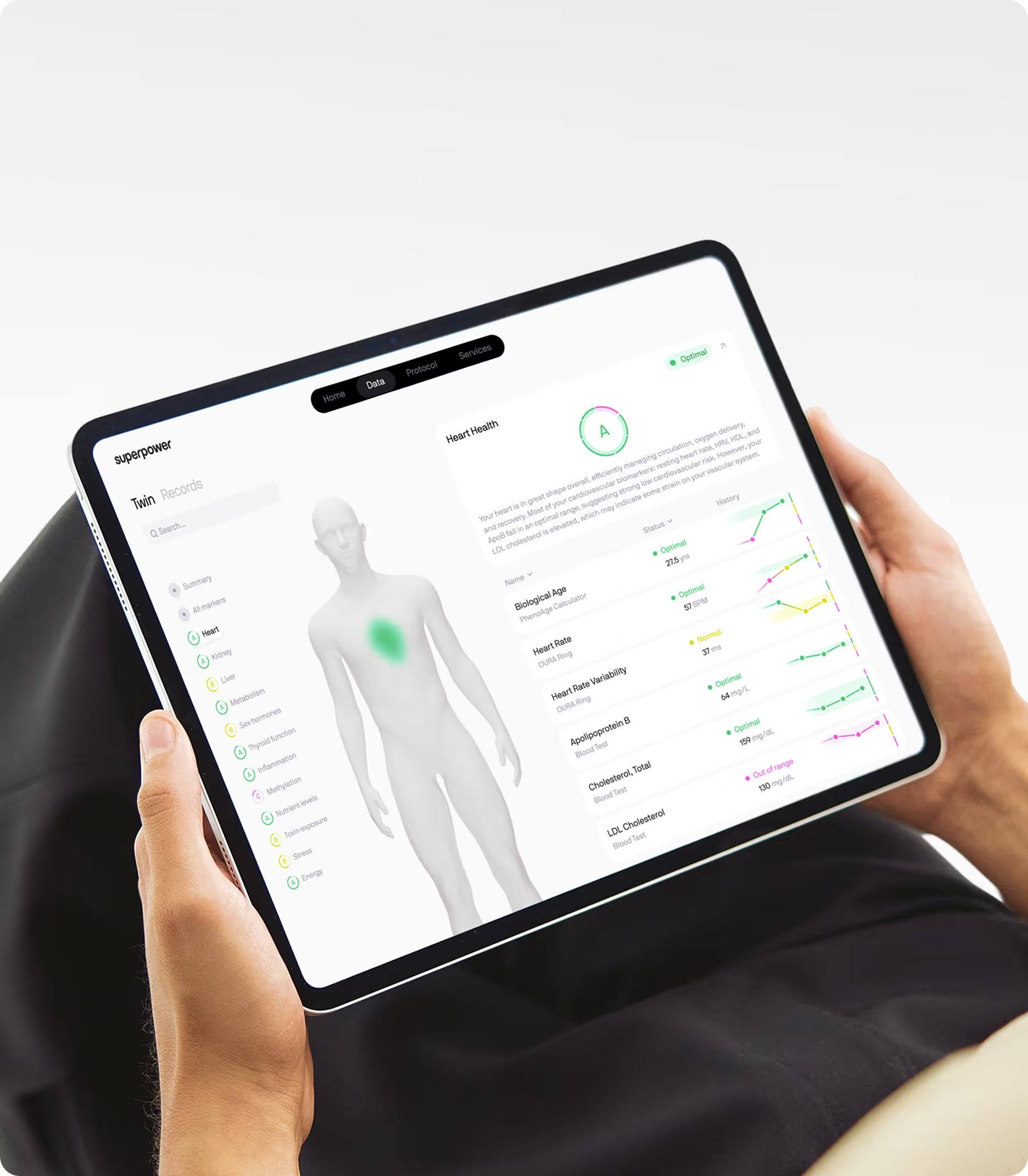
Measuring what actually matters for senescent cell burden
Tracking senescent cell accumulation and the effectiveness of senolytic interventions requires biomarkers that reflect cellular aging rather than just chronological age. Epigenetic clocks, particularly second-generation clocks like GrimAge and DunedinPACE, estimate biological age based on DNA methylation patterns and correlate with senescent cell burden. These are not direct measures of senescence, but they provide a longitudinal signal of aging rate that can be tracked over time.
Inflammatory markers offer another window:
- High-sensitivity C-reactive protein, IL-6, and TNF-alpha are components of the SASP and tend to be elevated in individuals with high senescent cell burdens.
- These markers are not specific to senescence, but a pattern of chronic low-grade elevation in the absence of acute infection or injury suggests inflammaging.
- Tracking these markers alongside metabolic health indicators like fasting insulin, HbA1c, and ApoB provides a more complete picture of how cellular aging intersects with metabolic and cardiovascular risk.
Body composition metrics are also relevant. Senescent cells accumulate disproportionately in adipose tissue, particularly visceral fat. DEXA scans that quantify visceral fat area and lean mass can provide indirect evidence of senescent cell burden, especially when tracked over time. Grip strength and VO2 max are functional markers that correlate with biological age and senescent cell load in animal models. These are not biomarkers of senescence per se, but they reflect the downstream consequences of cellular aging on physical resilience. For those considering senolytic interventions, baseline measurement of these markers before starting supplementation and repeat testing after several months can provide some indication of whether the intervention is moving the needle.
How to use longevity science to build a real baseline
If you want to understand how cellular aging is progressing in your own body, Superpower's 100+ biomarker panel covers the metabolic, inflammatory, and hormonal markers most relevant to senescent cell burden and biological aging. Tracking hsCRP, insulin, ApoB, homocysteine, and IGF-1 over time gives you a data-driven view of how well your metabolic and inflammatory systems are holding up as you age. These are the markers that standard annual bloodwork typically misses, but they're the ones that matter most for understanding whether interventions like dietary changes, exercise, or targeted supplementation are actually shifting your biological age trajectory. Longevity is not about chasing a single compound or protocol; it's about building a longitudinal picture of how your body is aging and using that data to make informed decisions about what to prioritize.



FAQs
Fisetin is a plant flavonoid classified as a senolytic, meaning it selectively induces death in senescent cells. It works by disrupting the BCL-2 family of anti-apoptotic proteins that senescent cells upregulate to resist clearance. By blocking these survival proteins, fisetin allows senescent cells to undergo apoptosis while leaving healthy proliferating cells largely unaffected. It also activates sirtuins, reduces oxidative stress via the NRF2 pathway, and influences mitochondrial health independently of its senolytic mechanism.
Strawberries contain the highest dietary concentration of fisetin at roughly 160 micrograms per gram of fresh weight. Other sources include apples, persimmons, onions, and cucumbers, but at substantially lower concentrations. A cup of strawberries provides approximately 0.16 milligrams of fisetin, while animal studies demonstrating lifespan extension use doses equivalent to roughly 7 grams for a 70 kg human per dose, making dietary intake alone insufficient to replicate research conditions.
Fisetin is a senolytic that eliminates existing senescent cells by lowering their apoptosis threshold. This differs mechanistically from senostatic agents like rapamycin, which suppress SASP secretion without killing senescent cells, and from interventions like caloric restriction, which reduce the rate of senescent cell formation. Combination senolytics like dasatinib plus quercetin also clear senescent cells but through partially different pathways; quercetin shows weaker solo senolytic activity than fisetin but is synergistic with dasatinib.
In aged mice, intermittent fisetin treatment extends median and maximum lifespan, reduces senescent cell markers in adipose tissue, kidney, and heart, and improves physical function. The doses used are typically 100 mg/kg body weight administered intermittently. Fisetin also shows tissue-specific variability, clearing senescent endothelial cells more effectively than fibroblasts in vitro, reflecting differences in BCL-2 family protein expression across cell types. Intermittent arterial function improvements have also been demonstrated in animal models.
Human trials are limited in number and scope. A 2019 pilot study tested fisetin at 100 mg twice daily for two days in women with obesity, finding the regimen safe and feasible with some reduction in inflammatory markers but no assessment of longevity outcomes. Ongoing trials are evaluating fisetin in osteoarthritis, chronic kidney disease, and frailty populations. Human evidence is primarily safety and feasibility data, not proof of lifespan or healthspan extension. Results from adequately powered longevity trials are not yet available.
Oral fisetin has poor absorption and rapid first-pass metabolism, resulting in low plasma concentrations after typical doses. A cup of strawberries provides about 0.16 milligrams, while research protocols use gram-level doses. Novel delivery systems using liposomal encapsulation or nanoparticle carriers are being developed to improve bioavailability, but these are not yet widely commercially available. The gut microbiome also influences flavonoid metabolism, creating inter-individual variability in systemic exposure from the same dose.
References
- Grynkiewicz, G., & Demchuk, O. M. (2019). New Perspectives for Fisetin. Frontiers in chemistry, 7, 697. https://doi.org/10.3389/fchem.2019.00697
- Yousefzadeh, M. J., Zhu, Y., McGowan, S. J., Angelini, L., Fuhrmann-Stroissnigg, H., Xu, M., Ling, Y. Y., Melos, K. I., Pirtskhalava, T., Inman, C. L., McGuckian, C., Wade, E. A., Kato, J. I., Grassi, D., Wentworth, M., Burd, C. E., Arriaga, E. A., Ladiges, W. L., Tchkonia, T., ... Niedernhofer, L. J. (2018). Fisetin is a senotherapeutic that extends health and lifespan. EBioMedicine, 36, 18-28. https://doi.org/10.1016/j.ebiom.2018.09.015
- Chaib, S., Tchkonia, T., & Kirkland, J. L. (2022). Cellular senescence and senolytics: the path to the clinic. Nature medicine, 28(8), 1556-1568. https://doi.org/10.1038/s41591-022-01923-y
- Mahoney, S. A., Venkatasubramanian, R., Darrah, M. A., Ludwig, K. R., VanDongen, N. S., Greenberg, N. T., Longtine, A. G., Hutton, D. A., Brunt, V. E., Campisi, J., Melov, S., Seals, D. R., Rossman, M. J., & Clayton, Z. S. (2024). Intermittent supplementation with fisetin improves arterial function in old mice by decreasing cellular senescence. Aging cell, 23(3), e14060. https://doi.org/10.1111/acel.14060
- Lee, E., & Burns, M. (2024). The Effects of Fisetin on Reducing Biological Aging: A Pilot Study. Alternative therapies in health and medicine, 30(9), 6-10. https://pubmed.ncbi.nlm.nih.gov/39269340/
























.avif)
.svg)
.svg)


